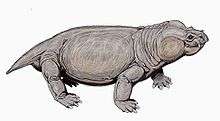
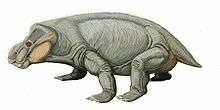
Anomodont
Anomodontia is an extinct group of non-mammalian therapsids containing many species from the Permian and Triassic periods (possibly continuing into the Early Cretaceous),[2] most of which were toothless, possibly endothermic[3][4][5] herbivores.[6] Anomodonts were very diverse during the Middle Permian, including primitive forms like Anomocephalus and Patranomodon and groups like Venyukovioidea, Dromasauria, and Dicynodontia. Of these, only the dicynodonts survived beyond the Middle Permian. Dicynodonts became the most successful and abundant of all herbivores in the Late Permian and Triassic, filling ecological niches ranging from large browsers down to small burrowers. Few dicynodont families survived the Permian–Triassic extinction event, but one lineage evolved into large, stocky forms that remained the dominant terrestrial herbivores right until the Late Triassic, when changing conditions caused them to decline.
Classification
Taxonomy
Phylogeny
Cladogram modified from Liu et al. (2009):[2]
Below is a cladogram from Kammerer et al. (2013).[7] The data matrix of Kammerer et al. (2013), a list of characteristics that was used in the analysis, was based on that of Kammerer et al. (2011), which followed a comprehensive taxonomic revision of Dicynodon.[8] Because of this, many of the relationships found by Kammerer et al. (2013) are the same as those found by Kammerer et al. (2011). However, several taxa were added to the analysis, including Tiarajudens Eubrachiosaurus, Shaanbeikannemeyeria, Zambiasaurus and many "outgroup" taxa (positioned outside Anomodontia), while other taxa were re-coded. As in Kammerer et al. (2011), the interrelationships of non-kannemeyeriiform dicynodontoids are weakly supported and thus vary between the analyses.[7]
| 1 |
1 Anomodontia, 2 Venyukovioidea, 3 Chainosauria, 4 Dicynodontia, 5 Therochelonia, 6 Diictodontia, 7 Pylaecephalidae, 8 Emydopoidea, 9 Kistecephalia, 10 Kingoriidae, 11 Cistecephalidae, 12 Bidentalia |
|
See also
References
- ↑
- Thulborn, T. & Turner, S. 2003. The last dicynodont: an Australian Cretaceous relict. Proceedings of the Royal Society of London B 270, 985-993. Abstract.
- 1 2 Liu, J.; Rubidge, B.; Li, J. (2009). "A new specimen of Biseridens qilianicus indicates its phylogenetic position as the most basal anomodont". Proceedings of the Royal Society B. 277 (1679): 285–292. doi:10.1098/rspb.2009.0883. PMC 2842672
 . PMID 19640887.
. PMID 19640887. - ↑ Bakker 1975
- ↑ JENNIFER BOTHA-BRINK and KENNETH D. ANGIELCZYK, Do extraordinarily high growth rates in Permo-Triassic dicynodonts (Therapsida, Anomodontia) explain their success before and after the end-Permian extinction?, Version of Record online: 26 JUL 2010 DOI: 10.1111/j.1096-3642.2009.00601.x
- ↑ Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia Piotr Bajdek1, Martin Qvarnström2, Krzysztof Owocki3, Tomasz Sulej3, Andrey G. Sennikov4,5, Valeriy K. Golubev4,5 andGrzegorz Niedźwiedzki2 Article first published online: 25 NOV 2015 DOI: 10.1111/let.12156
- ↑ Chinsamy-Turan, A. (2011) Forerunners of Mammals: Radiation - Histology - Biology, p.39. Indiana University Press, ISBN 0253356970. Retrieved May 2012
- 1 2 Kammerer, C. F.; Fröbisch, J. R.; Angielczyk, K. D. (2013). Farke, Andrew A, ed. "On the Validity and Phylogenetic Position of Eubrachiosaurus browni, a Kannemeyeriiform Dicynodont (Anomodontia) from Triassic North America". PLoS ONE. 8 (5): e64203. doi:10.1371/journal.pone.0064203.
- ↑ Kammerer, C.F.; Angielczyk, K.D.; Fröbisch, J. (2011). "A comprehensive taxonomic revision of Dicynodon (Therapsida, Anomodontia) and its implications for dicynodont phylogeny, biogeography, and biostratigraphy". Journal of Vertebrate Paleontology. 31 (Suppl. 1): 1–158. doi:10.1080/02724634.2011.627074.
|
|---|
|
|
|
|
|
|
|
|
|
|
|
| |
|---|
| Dinodontosauridae | |
|---|
| | |
|---|
| | |
|---|
| | |
|---|
| Other kannemeyeriforms | |
|---|
|
|
|
|