Polyomaviridae
| Polyomaviridae | |
|---|---|
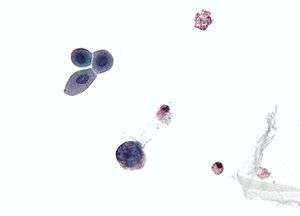 | |
| Micrograph showing a polyomavirus infected cell—large (blue) cell below-center-left. Urine cytology specimen. | |
| Virus classification | |
| Group: | Group I (dsDNA) |
| Family: | Polyomaviridae |
| Genera | |
Polyomaviridae is a family of viruses whose natural hosts are primarily mammals and birds. As of the most recent (2015) taxonomy release by the International Committee on Taxonomy of Viruses, there were 73 recognized species in this family contained within four genera, as well as three species that could not be assigned to a genus.[1] Of these, 13 species are known to infect humans.[2][3] Most of these viruses, such as BK virus and JC virus, are very common and typically asymptomatic in most human populations studied.[4][5] However, some polyomaviruses are associated with human disease, particularly in immunocompromised individuals; BK virus is associated with nephropathy in renal transplant and non-renal solid organ transplant patients,[6][7] JC virus with progressive multifocal leukoencephalopathy,[8] and Merkel cell virus with Merkel cell cancer.[9]
Some members of the family are oncoviruses, meaning they can cause tumors; they often persist as latent infections in a host without causing disease, but may produce tumors in a host of a different species, or in individuals with ineffective immune systems. The family was first discovered due to its oncogenic properties; some members of the family, most prominently murine polyomavirus, have been extensively studied in the laboratory to understand the mechanism by which they induce carcinogenesis. The name polyoma refers to the viruses' ability to produce multiple (poly-) tumors (-oma).
Structure and genome

Polyomaviruses are unenveloped double-stranded DNA viruses with circular genomes of around 5000 base pairs. The genome is packaged in a viral capsid of about 40-50 nanometers in diameter, which is icosahedral in shape (T=7 symmetry).[10] The capsid is composed of 72 pentamers of a protein called VP1, which is capable of self-assembly into a closed icosahedron;[11] each molecule of VP1 is associated with one molecule of one of the other two capsid proteins, VP2 or VP3.[12]
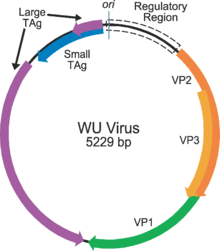
The genome of a typical polyomavirus codes for between 5 and 9 proteins, divided into two transcriptional regions called the early and late regions due to the time during infection in which they are transcribed. Each region is transcribed by the host cell's RNA polymerase II as a single pre-messenger RNA containing multiple genes. The early region usually codes for two proteins, the small and large tumor antigens, produced by alternative splicing. The late region contains the three capsid structural proteins VP1, VP2, and VP3, produced by alternative translational start sites. Additional genes and other variations on this theme are present in some viruses: for example, rodent polyomaviruses have a third protein called middle tumor antigen in the early region, which is extremely efficient at inducing cellular transformation; SV40 has an additional capsid protein VP4; some examples have an additional regulatory protein called agnoprotein expressed from the late region. The genome also contains a non-coding control or regulatory region containing the early and late regions' promoters, transcriptional start sites, and the origin of replication.[10][12]
| Genus | Structure | Symmetry | Capsid | Genomic arrangement | Genomic segmentation |
|---|---|---|---|---|---|
| Polyomavirus | Icosahedral | T=7 | Non-Enveloped | Circular | Monopartite |
Replication and life cycle
Prior to genome replication, the processes of viral attachment, entry and uncoating occur. Cellular receptors for polyomaviruses are sialic acid residues of glycans, commonly gangliosides. The attachment of polyomaviruses to host cells is mediated by the binding of VP1 to sialylated glycans on the cell surface.[14]
Polyomavirus virions are subsequently endocytosed and transported first to the endoplasmic reticulum where a conformational change occurs revealing Vp2. Then by an unknown mechanism the virus is exported to the nucleus.
Polyomaviruses replicate in the nucleus of the host. They are able to utilise the host’s machinery because the genomic structure is homologous to that of the mammalian host. Moreover, the promoter sequence of Polyomavirus' promoter sequence is a strong attractant for the host's RNAP. Viral replication occurs in two distinct phases; early and late gene expression, separated by genome replication.
Early gene expression is responsible for the synthesis of non-structural proteins. Since Polyomaviruses rely on the host to control both the gene expression, the role of the non-structural proteins is to regulate the cellular mechanisms. Close to the N terminal end of polyomavirus genome are enhancer elements which induce activation and transcription of a molecule known as the T-antigen (see SV40 large T-antigen). Early mRNA’s, encoding T-antigen are produced by host RNA polymerase II. T-antigen autoregulates early mRNA’s, subsequently leading to elevated levels of T-antigen. At high concentrations of T-antigen, early gene expression is repressed, triggering the late phase of viral infection to begin.
Genome replication acts to separate the early and late phase gene expression. The duplicated viral genome is synthesised and processed as if it were cellular DNA, exploiting the host’s machinery. As the daughter viral DNA are synthesised they associate with cellular nucleosomes to form structures that are often referred to as "minichromosomes". In this manner the DNA is packaged more efficiently.
Late gene expression synthesises the structural proteins, responsible for the viral particle composition. This occurs during and after genome replication. As with the early gene expression products, late gene expression generates an array of proteins as a result of alternative splicing.
Within each viral protein are 'nuclear localization signals' which cause the viral proteins to amass in the nucleus. Assembly of new virus particles consequently occurs within the nucleus of the host cell.[15]
Release of newly synthesized polyomavirus particles exit the infected cell by one of two mechanisms. Firstly and less commonly, they are transported in cytoplasmic vacuoles to the plasma membrane, where budding occurs. More frequently, they are released when the cell lyses due to the cytotoxicity of virus particles present in the infected cell.
| Genus | Host details | Tissue tropism | Entry details | Release details | Replication site | Assembly site | Transmission |
|---|---|---|---|---|---|---|---|
| Polyomavirus | Mammals; birds | Respiratory system; kidneys, brain | Cell receptor endocytosis | Lysis | Nucleus | Nucleus | Oral-fecal |
Viral proteins
The polyoma large and small T-antigens
The large T-antigen plays a key role in regulating the viral life cycle by binding to the viral origin of DNA replication where it promotes DNA synthesis. Also as the polyomavirus relies on the host cell machinery to replicate the host cell needs to be in s-phase for this to begin. Due to this, large T-antigen also modulates cellular signaling pathways to stimulate progression of the cell cycle by binding to a number of cellular control proteins.[16] This is achieved by a two prong attack of inhibiting tumor suppressing genes p53 and members of the retinoblastoma (pRB) family,[17] and stimulating cell growth pathways by binding cellular DNA, ATPase-helicase, DNA polymerase α association, and binding of transcription preinitiation complex factors.[18] This abnormal stimulation of the cell cycle is a powerful force for oncogenic transformation.
The small T-antigen protein is also able to activate several cellular pathways that stimulate cell proliferation. Polyomavirus small T antigens commonly target protein phosphatase 2A (PP2A),[19] a key multisubunit regulator of multiple pathways including Akt, the mitogen-activated protein kinase (MAPK) pathway, and the stress-activated protein kinase (SAPK) pathway.[20][21] Merkel cell polyomavirus small T antigen encodes a unique domain, called the LT-stabilization domain (LSD), that binds to and inhibits the FBXW7 E3 ligase regulating both cellular and viral oncoproteins.[22] Unlike for SV40, the MCV small T antigen directly transforms rodent cells in vitro.[23]
The polyoma middle T-antigen
The polyoma middle T-antigen is used in animal breast cancer model systems like the PYMT system where it is coupled to the MMTV promoter. There it functions as an oncogene, while the tissue where the tumor develops is determined by the MMTV promoter.
Agnoprotein
The agnoprotein is a small multifunctional phospho-protein found in the late coding part of the genome of some polyomaviruses, most notably BK virus, JC virus, and SV40. It is essential for proliferation in the viruses that express it and is thought to be involved in regulating the viral life cycle, particularly replication and viral exit from the host cell, but the exact mechanisms are unclear.[24][25]
Taxonomy
The polyomaviruses are members of group I (dsDNA viruses). The classification of Polyomaviruses has been the subject of several proposed revisions as new members of the group are discovered. Formerly, polyomaviruses and papillomaviruses, which share many structural features but have very different genomic organizations, were classified together in the now-obsolete family Papovaviridae.[1] (The name Papovaviridae derived from three abbreviations: Pa for Papillomavirus, Po for Polyomavirus, and Va for "vacuolating.")[26] The polyomaviruses were divided into three major clades (that is, genetically-related groups): the SV40 clade, the avian clade, and the murine polyomavirus clade.[27] A subsequent proposed reclassification by the International Committee on Taxonomy of Viruses (ICTV) recommended dividing the family of Polyomaviridae into three genera:[28]
- Genus Orthopolyomavirus (type species SV40)
- Genus Wukipolyomavirus (type species KI polyomavirus)
- Genus Avipolyomavirus (type species Avian polyomavirus)
The current ICTV classification system recognises four genera and 76 species, of which three could not be assigned a genus. This system retains the distinction between avian and mammalian viruses, grouping the avian subset into the genus Gammapolyomavirus.[1][2]
- Genus Alphapolyomavirus, type species Mus musculus polyomavirus 1 (murine polyomavirus)
- Genus Betapolyomavirus, type species Macaca mulatta polyomavirus 1 (SV40)
- Genus Gammapolyomavirus, type species Aves polyomavirus 1
- Genus Deltapolyomavirus, type species Human polyomavirus 6
Human polyomaviruses
Most polyomaviruses do not infect humans. Of the polyomaviruses cataloged as of 2016, a total of 13 were known with human hosts.[2] Many human polyomaviruses are very common and are asymptomatic.[4][5] However, some polyomaviruses are associated with human disease, particularly in immunocompromised individuals. MCV is highly divergent from the other human polyomaviruses and is most closely related to murine polyomavirus. Trichodysplasia spinulosa-associated polyomavirus (TSV) is distantly related to MCV. Two viruses—HPyV6 and HPyV7—are most closely related to KI and WU viruses, while HPyV9 is most closely related to the African green monkey-derived lymphotropic polyomavirus (LPV).
List of human polyomaviruses
The following 13 polyomaviruses with human hosts had been identified and had their genomes sequenced as of 2016:[2]
| Species | Proposed genus | Virus name | Abbreviation | NCBI RefSeq | Year of discovery | Clinical correlate (if any) | References |
|---|---|---|---|---|---|---|---|
| Human polyomavirus 5 | Alpha | Merkel cell polyomavirus | MCPyV | NC_010277 | 2008 | Merkel cell cancer[12] | [29][30][31] |
| Human polyomavirus 8 | Alpha | Trichodysplasia spinulosa polyomavirus | TSPyV | NC_014361 | 2010 | Trichodysplasia spinulosa[12] | [32][33] |
| Human polyomavirus 9 | Alpha | Human polyomavirus 9 | HPyV9 | NC_015150 | 2011 | None known | [34] |
| Human polyomavirus 12 | Alpha | Human polyomavirus 12 | HPyV12 | NC_020890 | 2013 | None known | [35] |
| Human polyomavirus 13 | Alpha | New Jersey polyomavirus | NJPyV | NC_024118 | 2014 | None known | [36] |
| Human polyomavirus 1 | Beta | BK polyomavirus | BKPyV | NC_001538 | 1971 | Polyomavirus-associated nephropathy; haemorrhagic cystitis[12] | [37] |
| Human polyomavirus 2 | Beta | JC polyomavirus | JCPyV | NC_001699 | 1971 | Progressive multifocal leukoencephalopathy[12] | [38] |
| Human polyomavirus 3 | Beta | KI polyomavirus | KIPyV | NC_009238 | 2007 | None known | [39] |
| Human polyomavirus 4 | Beta | WU polyomavirus | WUPyV | NC_009539 | 2007 | None known | [40] |
| Human polyomavirus 6 | Delta | Human polyomavirus 6 | HPyV6 | NC_014406 | 2010 | None known | [41] |
| Human polyomavirus 7 | Delta | Human polyomavirus 7 | HPyV7 | NC_014407 | 2010 | HPyV7-related epithelial hyperplasia[42][43] | [41] |
| Human polyomavirus 10 | Delta | MW polyomavirus | MWPyV | NC_018102 | 2012 | None known | [44][45][46] |
| Human polyomavirus 11 | Delta | STL polyomavirus | SLTPyV | NC_020106 | 2013 | None known | [47] |
The proposed Deltapolyomavirus genus contains only the four human viruses shown, with human polyomavirus 6 as the type species. The Alpha and Beta groups contain viruses that infect a variety of mammals. The Gamma group contains the avian viruses.[2] Clinically significant disease associations are shown only where causality is expected.[12][48]
Clinical relevance
All the polyomaviruses are highly common childhood and young adult infections.[49] Most of these infections appear to cause little or no symptoms. These viruses are probably lifelong persistent among almost all adults. Diseases caused by human polyomavirus infections are most common among immunocompromised people; disease associations include BK virus with nephropathy in renal transplant and non-renal solid organ transplant patients,[6][7] JC virus with progressive multifocal leukoencephalopathy,[8] and Merkel cell virus (MCV) with Merkel cell cancer.[9]
SV40
SV40 replicates in the kidneys of monkeys without causing disease, but can cause cancer in rodents under laboratory conditions. In the 1950s and early 1960s, well over 100 million people may have been exposed to SV40 due to previously undetected SV40 contamination of polio vaccine, prompting concern about the possibility that the virus might cause disease in humans.[50][51] Although it has been reported as present in some human cancers, including brain tumors, bone tumors, mesotheliomas, and non-Hodgkin's lymphomas,[52] accurate detection is often confounded by high levels of cross-reactivity for SV40 with widespread human polyomaviruses.[51] Most virologists dismiss SV40 as a cause for human cancers.[50][53][54]
Diagnosis
The diagnosis of polyomavirus almost always occurs after the primary infection as it is either asymptomatic or sub-clinical. Antibody assays are commonly used to detect presence of antibodies against individual viruses.[55] Competition assays are frequently needed to distinguish among highly similar polyomaviruses.[56]
In cases of progressive multifocal leucoencephalopathy (PML), a cross-reactive antibody to SV40 T antigen (commonly Pab419) is used to stain tissues directly for the presence of JC virus T antigen. PCR can be used on a biopsy of the tissue or cerebrospinal fluid to amplify the polyomavirus DNA. This allows not only the detection of polyomavirus but also which sub type it is.[57]
There are three main diagnostic techniques used for the diagnosis of the reactivation of polyomavirus in polyomavirus nephropathy (PVN): urine cytology, quantification of the viral load in both urine and blood, and a renal biopsy.[55] The reactivation of polyomavirus in the kidneys and urinary tract causes the shedding of infected cells, virions, and/or viral proteins in the urine. This allows urine cytology to examine these cells, which if there is polyomavirus inclusion of the nucleus, is diagnostic of infection.[58] Also as the urine of an infected individual will contain virions and/or viral DNA, quanitation of the viral load can be done through PCR.[59] This is also true for the blood.
Renal biopsy can also be used if the two methods just described are inconclusive or if the specific viral load for the renal tissue is desired. Similarly to the urine cytology, the renal cells are examined under light microscopy for polyomavirus inclusion of the nucleus, as well as cell lysis and viral partials in the extra cellular fluid. The viral load as before is also measure by PCR.
Tissue staining using a monoclonal antibody against MCV T antigen shows utility in differentiating Merkel cell carcinoma from other small, round cell tumors.[60] Blood tests to detect MCV antibodies have been developed and show that infection with the virus is widespread although Merkel cell carcinoma patients have exceptionally higher antibody responses than asymptomatically infected persons.[61][62][63][64]
History
Murine polyomavirus was the first polyomavirus discovered, having been reported by Ludwik Gross in 1953 as an extract of mouse leukemias capable of inducing parotid gland tumors.[65] The causative agent was identified as a virus by Sarah Stewart and Bernice Eddy, after whom it was once called "SE polyoma".[66][67][68] The term "polyoma" refers to the viruses' ability to produce multiple (poly-) tumors (-oma) under certain conditions. The name has been criticized as a "meatless linguistic sandwich" ("meatless" because both morphemes in "polyoma" are affixes) giving little insight into the viruses' biology; in fact, subsequent research has found that most polyomaviruses rarely cause clinically significant disease in their host organisms under natural conditions.[69]
Dozens of polyomaviruses have been identified and sequenced as of 2016, infecting mainly birds and mammals. Two polyomaviruses are known to infect fish, the black sea bass[70] and gilthead seabream.[71] A total of thirteen polyomaviruses are known to infect humans.[2]
References
- 1 2 3 ICTV. "Virus Taxonomy: 2015 Release". Retrieved 26 July 2016.
- 1 2 3 4 5 6 Polyomaviridae Study Group of the International Committee on Taxonomy of, Viruses; Calvignac-Spencer, S; Feltkamp, MC; Daugherty, MD; Moens, U; Ramqvist, T; Johne, R; Ehlers, B (29 February 2016). "A taxonomy update for the family Polyomaviridae.". Archives of Virology. 161: 1739–50. doi:10.1007/s00705-016-2794-y. PMID 26923930.
- ↑ DeCaprio, JA; Garcea, RL (2013). "A cornucopia of human polyomaviruses". Nat. Rev. Microbiol. 11: 264–76. doi:10.1038/nrmicro2992. PMC 3928796
 . PMID 23474680.
. PMID 23474680. - 1 2 Gossai, A; Waterboer, T; Nelson, HH; Michel, A; Willhauck-Fleckenstein, M; Farzan, SF; Hoen, AG; Christensen, BC; Kelsey, KT; Marsit, CJ; Pawlita, M; Karagas, MR (1 January 2016). "Seroepidemiology of Human Polyomaviruses in a US Population.". American Journal of Epidemiology. 183 (1): 61–9. doi:10.1093/aje/kwv155. PMID 26667254.
- 1 2 Kean, JM; Rao, S; Wang, M; Garcea, RL (March 2009). "Seroepidemiology of human polyomaviruses.". PLOS Pathogens. 5 (3): e1000363. doi:10.1371/journal.ppat.1000363. PMC 2655709. PMID 19325891.
- 1 2 Jamboti, JS (18 January 2016). "BK virus nephropathy in renal transplant recipients.". Nephrology (Carlton, Vic.). 21: 647–54. doi:10.1111/nep.12728. PMID 26780694.
- 1 2 Kuppachi, S; Kaur, D; Holanda, DG; Thomas, CP (April 2016). "BK polyoma virus infection and renal disease in non-renal solid organ transplantation.". Clinical kidney journal. 9 (2): 310–8. doi:10.1093/ckj/sfv143. PMC 4792618. PMID 26985385.
- 1 2 Adang, L; Berger, J (2015). "Progressive Multifocal Leukoencephalopathy.". F1000Research. 4. doi:10.12688/f1000research.7071.1. PMC 4754031. PMID 26918152.
- 1 2 Feng, H.; Shuda, M.; Chang, Y.; Moore, P. S. (2008). "Clonal Integration of a Polyomavirus in Human Merkel Cell Carcinoma". Science. 319 (5866): 1096–100. doi:10.1126/science.1152586. PMC 2740911. PMID 18202256.
- 1 2 "Viral Zone". ExPASy. Retrieved 15 June 2015.
- ↑ Salunke, DM; Caspar, DL; Garcea, RL (12 September 1986). "Self-assembly of purified polyomavirus capsid protein VP1.". Cell. 46 (6): 895–904. doi:10.1016/0092-8674(86)90071-1. PMID 3019556.
- 1 2 3 4 5 6 7 DeCaprio, JA; Garcea, RL (April 2013). "A cornucopia of human polyomaviruses.". Nature reviews. Microbiology. 11 (4): 264–76. doi:10.1038/nrmicro2992. PMC 3928796. PMID 23474680.
- ↑ Gaynor, AM; Nissen, MD; Whiley, DM; Mackay, IM; Lambert, SB; Wu, G; Brennan, DC; Storch, GA; Sloots, TP; Wang, D (4 May 2007). "Identification of a novel polyomavirus from patients with acute respiratory tract infections.". PLOS Pathogens. 3 (5): e64. doi:10.1371/journal.ppat.0030064. PMC 1864993. PMID 17480120.
- 1 2 Buch, MH; Liaci, AM; O'Hara, SD; Garcea, RL; Neu, U; Stehle, T (October 2015). "Structural and Functional Analysis of Murine Polyomavirus Capsid Proteins Establish the Determinants of Ligand Recognition and Pathogenicity.". PLOS Pathogens. 11 (10): e1005104. doi:10.1371/journal.ppat.1005104. PMC 4608799. PMID 26474293.
- ↑ Kazem, Siamaque; Van Der Meijden, Els; Kooijman, Sander; Rosenberg, Arlene S.; Hughey, Lauren C.; Browning, John C.; Sadler, Genevieve; Busam, Klaus; Pope, Elena; Benoit, Taylor; Fleckman, Philip; De Vries, Esther; Eekhof, Just A.; Feltkamp, Mariet C.W. (2012). "Trichodysplasia spinulosa is characterized by active polyomavirus infection". Journal of Clinical Virology. 53 (3): 225–30. doi:10.1016/j.jcv.2011.11.007. PMID 22196870.
- ↑ White, Martyn K.; Gordon, Jennifer; Reiss, Krzysztof; Del Valle, Luis; Croul, Sidney; Giordano, Antonio; Darbinyan, Armine; Khalili, Kamel (2005). "Human polyomaviruses and brain tumors". Brain Research Reviews. 50 (1): 69–85. doi:10.1016/j.brainresrev.2005.04.007. PMID 15982744.
- ↑ Kazem, Siamaque; Van Der Meijden, Els; Wang, Richard C.; Rosenberg, Arlene S.; Pope, Elena; Benoit, Taylor; Fleckman, Philip; Feltkamp, Mariet C. W. (2014). "Polyomavirus-Associated Trichodysplasia Spinulosa Involves Hyperproliferation, pRB Phosphorylation and Upregulation of p16 and p21". PLoS ONE. 9 (10): e108947. doi:10.1371/journal.pone.0108947. PMC 4188587. PMID 25291363.
- ↑ Kelley, W. L.; Georgopoulos, C (1997). "The T/t common exon of simian virus 40, JC, and BK polyomavirus T antigens can functionally replace the J-domain of the Escherichia coli DnaJ molecular chaperone". Proceedings of the National Academy of Sciences of the United States of America. 94 (8): 3679–3684. Bibcode:1997PNAS...94.3679K. doi:10.1073/pnas.94.8.3679. PMC 20500. PMID 9108037.
- ↑ Pallas, D. C.; Shahrik, L. K.; Martin, B. L.; Jaspers, S; Miller, T. B.; Brautigan, D. L.; Roberts, T. M. (1990). "Polyoma small and middle T antigens and SV40 small t antigen form stable complexes with protein phosphatase 2A". Cell. 60 (1): 167–76. doi:10.1016/0092-8674(90)90726-u. PMID 2153055.
- ↑ Sontag, Estelle; Fedorov, Sergei; Kamibayashi, Craig; Robbins, David; Cobb, Melanie; Mumby, Marc (1993). "The interaction of SV40 small tumor antigen with protein phosphatase 2A stimulates the map kinase pathway and induces cell proliferation". Cell. 75 (5): 887–97. doi:10.1016/0092-8674(93)90533-V. PMID 8252625.
- ↑ Watanabe, G; Howe, A; Lee, R. J.; Albanese, C; Shu, I. W.; Karnezis, A. N.; Zon, L; Kyriakis, J; Rundell, K; Pestell, R. G. (1996). "Induction of cyclin D1 by simian virus 40 small tumor antigen". Proceedings of the National Academy of Sciences of the United States of America. 93 (23): 12861–12866. Bibcode:1996PNAS...9312861W. doi:10.1073/pnas.93.23.12861. PMC 24011. PMID 8917510.
- ↑ Kwun, Hyun Jin; Shuda, Masahiro; Feng, Huichen; Camacho, Carlos J.; Moore, Patrick S.; Chang, Yuan (2013). "Merkel Cell Polyomavirus Small T Antigen Controls Viral Replication and Oncoprotein Expression by Targeting the Cellular Ubiquitin Ligase SCFFbw7". Cell Host & Microbe. 14 (2): 125–135. doi:10.1016/j.chom.2013.06.008.
- ↑ Shuda, Masahiro; Kwun, Hyun Jin; Feng, Huichen; Chang, Yuan; Moore, Patrick S. (2011). "Human Merkel cell polyomavirus small T antigen is an oncoprotein targeting the 4E-BP1 translation regulator". Journal of Clinical Investigation. 121 (9): 3623–34. doi:10.1172/JCI46323. PMID 21841310.
- ↑ Sariyer, Ilker K; Saribas, Abdullah S; White, Martyn K; Safak, Mahmut (2011). "Infection by agnoprotein-negative mutants of polyomavirus JC and SV40 results in the release of virions that are mostly deficient in DNA content". Virology Journal. 8: 255. doi:10.1186/1743-422X-8-255. PMC 3127838. PMID 21609431.
- ↑ Saribas, AS; Coric, P; Hamazaspyan, A; Davis, W; Axman, R; White, MK; Abou-Gharbia, M; Childers, W; Condra, JH; Bouaziz, S; Safak, M (October 2016). "Emerging From the Unknown: Structural and Functional Features of Agnoprotein of Polyomaviruses.". Journal of cellular physiology. 231 (10): 2115–27. doi:10.1002/jcp.25329. PMID 26831433.
- ↑ International Agency for Research on Cancer (2013). "IARC Working Group on the Evaluation of Carcinogenic Risk to Humans. Malaria and Some Polyomaviruses (SV40, BK, JC, and Merkel Cell Viruses).". IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. 104.
- ↑ Perez-Losada, M.; Christensen, R. G.; McClellan, D. A.; Adams, B. J.; Viscidi, R. P.; Demma, J. C.; Crandall, K. A. (2006). "Comparing Phylogenetic Codivergence between Polyomaviruses and Their Hosts". Journal of Virology. 80 (12): 5663–9. doi:10.1128/JVI.00056-06. PMC 1472594. PMID 16731904.
- ↑ Johne, Reimar; Buck, Christopher B.; Allander, Tobias; Atwood, Walter J.; Garcea, Robert L.; Imperiale, Michael J.; Major, Eugene O.; Ramqvist, Torbjorn; Norkin, Leonard C. (2011). "Taxonomical developments in the family Polyomaviridae". Archives of Virology. 156 (9): 1627–34. doi:10.1007/s00705-011-1008-x. PMC 3815707. PMID 21562881.
- ↑ Altman, Lawreence K. (2008-01-18). "Virus Is Linked to a Powerful Skin Cancer". The New York Times. Retrieved 2008-01-18.
- ↑ Feng, H.; Shuda, M.; Chang, Y.; Moore, P. S. (2008). "Clonal Integration of a Polyomavirus in Human Merkel Cell Carcinoma". Science. 319 (5866): 1096–100. Bibcode:2008Sci...319.1096F. doi:10.1126/science.1152586. PMC 2740911. PMID 18202256.
- ↑ Shuda, Masahiro; Arora, Reety; Kwun, Hyun Jin; Feng, Huichen; Sarid, Ronit; Fernández-Figueras, María-Teresa; Tolstov, Yanis; Gjoerup, Ole; Mansukhani, Mahesh M.; Swerdlow, Steven H.; Chaudhary, Preet M.; Kirkwood, John M.; Nalesnik, Michael A.; Kant, Jeffrey A.; Weiss, Lawrence M.; Moore, Patrick S.; Chang, Yuan (2009). "Human Merkel cell polyomavirus infection I. MCV T antigen expression in Merkel cell carcinoma, lymphoid tissues and lymphoid tumors". International Journal of Cancer. 125 (6): 1243–9. doi:10.1002/ijc.24510. PMID 19499546.
- ↑ Van Der Meijden, Els; Janssens, René W. A.; Lauber, Chris; Bouwes Bavinck, Jan Nico; Gorbalenya, Alexander E.; Feltkamp, Mariet C. W. (2010). "Discovery of a New Human Polyomavirus Associated with Trichodysplasia Spinulosa in an Immunocompromized Patient". PLoS Pathogens. 6 (7): e1001024. doi:10.1371/journal.ppat.1001024. PMC 2912394. PMID 20686659.
- ↑ Kazem, S; van der Meijden, E; Feltkamp, MC (August 2013). "The trichodysplasia spinulosa-associated polyomavirus: virological background and clinical implications.". APMIS : acta pathologica, microbiologica, et immunologica Scandinavica. 121 (8): 770–82. doi:10.1111/apm.12092. PMID 23593936.
- ↑ Scuda, N; Hofmann, J; Calvignac-Spencer, S; Ruprecht, K; Liman, P; Kühn, J; Hengel, H; Ehlers, B (May 2011). "A novel human polyomavirus closely related to the african green monkey-derived lymphotropic polyomavirus.". Journal of Virology. 85 (9): 4586–90. doi:10.1128/jvi.02602-10. PMC 3126223. PMID 21307194.
- ↑ Korup, Sarah; Rietscher, Janita; Calvignac-Spencer, Sébastien; Trusch, Franziska; Hofmann, Jörg; Moens, Ugo; Sauer, Igor; Voigt, Sebastian; Schmuck, Rosa; Ehlers, Bernhard (2013). "Identification of a Novel Human Polyomavirus in Organs of the Gastrointestinal Tract". PLoS ONE. 8 (3): e58021. Bibcode:2013PLoSO...858021K. doi:10.1371/journal.pone.0058021. PMC 3596337. PMID 23516426.
- ↑ Mishra, N; Pereira, M; Rhodes, RH; An, P; Pipas, JM; Jain, K; Kapoor, A; Briese, T; Faust, PL; Lipkin, WI (15 November 2014). "Identification of a novel polyomavirus in a pancreatic transplant recipient with retinal blindness and vasculitic myopathy.". The Journal of Infectious Diseases. 210 (10): 1595–9. doi:10.1093/infdis/jiu250. PMC 4334791. PMID 24795478.
- ↑ Gardner, SD; Field, AM; Coleman, DV; Hulme, B (19 June 1971). "New human papovavirus (B.K.) isolated from urine after renal transplantation.". Lancet (London, England). 1 (7712): 1253–7. doi:10.1016/s0140-6736(71)91776-4. PMID 4104714.
- ↑ Padgett, BL; Walker, DL; ZuRhein, GM; Eckroade, RJ; Dessel, BH (19 June 1971). "Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy.". Lancet (London, England). 1 (7712): 1257–60. doi:10.1016/S0140-6736(71)91777-6. PMID 4104715.
- ↑ Allander, T.; Andreasson, K.; Gupta, S.; Bjerkner, A.; Bogdanovic, G.; Persson, M. A. A.; Dalianis, T.; Ramqvist, T.; Andersson, B. (2007). "Identification of a Third Human Polyomavirus". Journal of Virology. 81 (8): 4130–6. doi:10.1128/JVI.00028-07. PMC 1866148. PMID 17287263.
- ↑ Gaynor, Anne M.; Nissen, Michael D.; Whiley, David M.; MacKay, Ian M.; Lambert, Stephen B.; Wu, Guang; Brennan, Daniel C.; Storch, Gregory A.; Sloots, Theo P.; Wang, David (2007). "Identification of a Novel Polyomavirus from Patients with Acute Respiratory Tract Infections". PLoS Pathogens. 3 (5): e64. doi:10.1371/journal.ppat.0030064. PMC 1864993. PMID 17480120.
- 1 2 Schowalter, Rachel M.; Pastrana, Diana V.; Pumphrey, Katherine A.; Moyer, Adam L.; Buck, Christopher B. (2010). "Merkel Cell Polyomavirus and Two Previously Unknown Polyomaviruses Are Chronically Shed from Human Skin". Cell Host & Microbe. 7 (6): 509–15. doi:10.1016/j.chom.2010.05.006. PMC 2919322. PMID 20542254.
- ↑ Ho, J; Jedrych, JJ; Feng, H; Natalie, AA; Grandinetti, L; Mirvish, E; Crespo, MM; Yadav, D; Fasanella, KE; Proksell, S; Kuan, SF; Pastrana, DV; Buck, CB; Shuda, Y; Moore, PS; Chang, Y (15 May 2015). "Human polyomavirus 7-associated pruritic rash and viremia in transplant recipients.". The Journal of Infectious Diseases. 211 (10): 1560–5. doi:10.1093/infdis/jiu524. PMC 4425822. PMID 25231015.
- ↑ Toptan, Tuna; Yousem, Samuel A.; Ho, Jonhan; Matsushima, Yuki; Stabile, Laura P.; Fernández-Figueras, Maria-Teresa; Bhargava, Rohit; Ryo, Akihide; Moore, Patrick S.; Chang, Yuan (25 February 2016). "Survey for human polyomaviruses in cancer". JCI Insight. 1 (2). doi:10.1172/jci.insight.85562. PMC 4811373. PMID 27034991.
- ↑ Siebrasse, E. A.; Reyes, A.; Lim, E. S.; Zhao, G.; Mkakosya, R. S.; Manary, M. J.; Gordon, J. I.; Wang, D. (2012). "Identification of MW Polyomavirus, a Novel Polyomavirus in Human Stool". Journal of Virology. 86 (19): 10321–6. doi:10.1128/JVI.01210-12. PMID 22740408.
- ↑ Buck, C. B.; Phan, G. Q.; Raiji, M. T.; Murphy, P. M.; McDermott, D. H.; McBride, A. A. (2012). "Complete Genome Sequence of a Tenth Human Polyomavirus". Journal of Virology. 86 (19): 10887. doi:10.1128/JVI.01690-12. PMID 22966183.
- ↑ Yu, Guixia; Greninger, Alexander L.; Isa, Pavel; Phan, Tung G.; Martínez, Miguel Angel; de la Luz Sanchez, Maria; Contreras, Juan Francisco; Santos-Preciado, José Ignacio; Parsonnet, Julie; Miller, Steve; Derisi, Joseph L.; Delwart, Eric; Arias, Carlos F.; Chiu, Charles Y. (2012). "Discovery of a Novel Polyomavirus in Acute Diarrheal Samples from Children". PLoS ONE. 7 (11): e49449. Bibcode:2012PLoSO...749449Y. doi:10.1371/journal.pone.0049449. PMC 3498111. PMID 23166671.
- ↑ Lim, Efrem S.; Reyes, Alejandro; Antonio, Martin; Saha, Debasish; Ikumapayi, Usman N.; Adeyemi, Mitchell; Stine, O. Colin; Skelton, Rebecca; Brennan, Daniel C.; Mkakosya, Rajhab S.; Manary, Mark J.; Gordon, Jeffrey I.; Wang, David (2013). "Discovery of STL polyomavirus, a polyomavirus of ancestral recombinant origin that encodes a unique T antigen by alternative splicing". Virology. 436 (2): 295–303. doi:10.1016/j.virol.2012.12.005. PMID 23276405.
- ↑ Dalianis, T; Hirsch, HH (15 March 2013). "Human polyomaviruses in disease and cancer.". Virology. 437 (2): 63–72. doi:10.1016/j.virol.2012.12.015. PMID 23357733.
- ↑ Egli, Adrian; Infanti, Laura; Dumoulin, Alexis; Buser, Andreas; Samaridis, Jacqueline; Stebler, Christine; Gosert, Rainer; Hirsch, Hans H. (2009). "Prevalence of Polyomavirus BK and JC Infection and Replication in 400 Healthy Blood Donors". The Journal of Infectious Diseases. 199 (6): 837–46. doi:10.1086/597126. PMID 19434930.
- 1 2 Poulin, D. L.; Decaprio, J. A. (2006). "Is There a Role for SV40 in Human Cancer?". Journal of Clinical Oncology. 24 (26): 4356–65. doi:10.1200/JCO.2005.03.7101. PMID 16963733.
- 1 2 Zur Hausen, Harald (2003). "Sv40 in human cancers—An endless tale?". International Journal of Cancer. 107 (5): 687. doi:10.1002/ijc.11517. PMID 14566815.
- ↑ Gazdar, AF; Butel, JS; Carbone, M (December 2002). "SV40 and human tumours: myth, association or causality?". Nature reviews. Cancer. 2 (12): 957–64. doi:10.1038/nrc947. PMID 12459734.
- ↑ Carroll-Pankhurst, C; Engels, EA; Strickler, HD; Goedert, JJ; Wagner, J; Mortimer EA Jr. (Nov 2001). "Thirty-five year mortality following receipt of SV40- contaminated polio vaccine during the neonatal period.". Br J Cancer. 85 (9): 1295–7. doi:10.1054/bjoc.2001.2065. PMID 11720463.
- ↑ Shah, KV (15 January 2007). "SV40 and human cancer: a review of recent data.". International Journal of Cancer. 120 (2): 215–23. doi:10.1002/ijc.22425. PMID 17131333.
- 1 2 Drachenberg, Cinthia B.; Hirsch, Hans H.; Ramos, Emilio; Papadimitriou, John C. (2005). "Polyomavirus disease in renal transplantation: Review of pathological findings and diagnostic methods". Human Pathology. 36 (12): 1245–55. doi:10.1016/j.humpath.2005.08.009. PMID 16311117.
- ↑ Viscidi, Raphael P.; Clayman, Barbara (2006). "Serological Cross Reactivity between Polyomavirus Capsids". In Ahsan, Nasimul. Polyomaviruses and Human Diseases. Advances in Experimental Medicine and Biology. 577. pp. 73–84. doi:10.1007/0-387-32957-9_5. ISBN 978-0-387-29233-5. PMID 16626028.
- ↑ Drews, Klaus; Bashir, Tarig; Dörries, Kristina (2000). "Quantification of human polyomavirus JC in brain tissue and cerebrospinal fluid of patients with progressive multifocal leukoencephalopathy by competitive PCR". Journal of Virological Methods. 84 (1): 23–36. doi:10.1016/S0166-0934(99)00128-7. PMID 10644084.
- ↑ Nickeleit, V; Hirsch, H. H.; Binet, I. F.; Gudat, F; Prince, O; Dalquen, P; Thiel, G; Mihatsch, M. J. (May 1999). "Polyomavirus infection of renal allograft recipients: from latent infection to manifest disease". Journal of the American Society of Nephrology. 10 (5): 1080–9. PMID 10232695.
- ↑ Randhawa, Parmjeet S.; Vats, Abhay; Zygmunt, Deborah; Swalsky, Patricia; Scantlebury, Velma; Shapiro, Ron; Finkelstein, Sydney (2002). "Quantitation of viral DNA in renal allograft tissue from patients with BK virus nephropathy1". Transplantation. 74 (4): 485–8. doi:10.1097/00007890-200208270-00009. PMID 12352906.
- ↑ Busam, K. J.; Jungbluth, A. A.; Rekthman, N; Coit, D; Pulitzer, M; Bini, J; Arora, R; Hanson, N. C.; Tassello, J. A.; Frosina, D; Moore, P; Chang, Y (2009). "Merkel Cell Polyomavirus Expression in Merkel Cell Carcinomas and Its Absence in Combined Tumors and Pulmonary Neuroendocrine Carcinomas". The American journal of surgical pathology. 33 (9): 1378–1385. doi:10.1097/PAS.0b013e3181aa30a5. PMC 2932664. PMID 19609205.
- ↑ Kean, J. M.; Rao, S; Wang, M; Garcea, R. L. (2009). "Seroepidemiology of Human Polyomaviruses". PLoS Pathogens. 5 (3): e1000363. doi:10.1371/journal.ppat.1000363. PMC 2655709. PMID 19325891.
- ↑ Tolstov, Y. L.; Pastrana, D. V.; Feng, H; Becker, J. C.; Jenkins, F. J.; Moschos, S; Chang, Y; Buck, C. B.; Moore, P. S. (2009). "Human Merkel cell polyomavirus infection II. MCV is a common human infection that can be detected by conformational capsid epitope immunoassays". International Journal of Cancer. Journal International Du Cancer. 125 (6): 1250–1256. doi:10.1002/ijc.24509. PMC 2747737. PMID 19499548.
- ↑ Pastrana, D. V.; Tolstov, Y. L.; Becker, J. C.; Moore, P. S.; Chang, Y; Buck, C. B. (2009). "Quantitation of Human Seroresponsiveness to Merkel Cell Polyomavirus". PLoS Pathogens. 5 (9): e1000578. doi:10.1371/journal.ppat.1000578. PMC 2734180. PMID 19750217.
- ↑ Carter, J. J.; Paulson, K. G.; Wipf, G. C.; Miranda, D; Madeleine, M. M.; Johnson, L. G.; Lemos, B. D.; Lee, S; Warcola, A. H.; Iyer, J. G.; Nghiem, P; Galloway, D. A. (2009). "Association of Merkel Cell Polyomavirus–Specific Antibodies with Merkel Cell Carcinoma". JNCI Journal of the National Cancer Institute. 101 (21): 1510–1522. doi:10.1093/jnci/djp332. PMC 2773184. PMID 19776382.
- ↑ Gross, L. (1953). "A Filterable Agent, Recovered from Ak Leukemic Extracts, Causing Salivary Gland Carcinomas in C3H Mice". Experimental Biology and Medicine. 83 (2): 414–21. doi:10.3181/00379727-83-20376. PMID 13064287.
- ↑ STEWART, SE; EDDY, BE; BORGESE, N (June 1958). "Neoplasms in mice inoculated with a tumor agent carried in tissue culture.". Journal of the National Cancer Institute. 20 (6): 1223–43. PMID 13549981.
- ↑ Eddy, Bernice E.; Stewart, Sarah E. (November 1959). "Characteristics of the SE Polyoma Virus". American Journal of Public Health and the Nations Health. 49 (11): 1486–1492. doi:10.2105/AJPH.49.11.1486.
- ↑ Percy, Dean H.; Barthold, Stephen W. (2013). "Polyoma Virus Infection". Pathology of Laboratory Rodents and Rabbits (3rd ed.). John Wiley & Sons. ISBN 1118704630.
- ↑ Gottlieb, KA; Villarreal, LP (June 2001). "Natural biology of polyomavirus middle T antigen.". Microbiology and molecular biology reviews : MMBR. 65 (2): 288–318 ; second and third pages, table of contents. doi:10.1128/mmbr.65.2.288-318.2001. PMC 99028. PMID 11381103.
- ↑ Peretti, A; FitzGerald, PC; Bliskovsky, V; Pastrana, DV; Buck, CB (29 January 2015). "Genome Sequence of a Fish-Associated Polyomavirus, Black Sea Bass (Centropristis striata) Polyomavirus 1.". Genome announcements. 3 (1): e01476–14. doi:10.1128/genomeA.01476-14. PMC 4319505. PMID 25635011.
- ↑ López-Bueno, A; Mavian, C; Labella, AM; Castro, D; Borrego, JJ; Alcami, A; Alejo, A (20 July 2016). "Concurrence of iridovirus, polyomavirus and a unique member of a new group of fish papillomaviruses in lymphocystis disease affected gilthead seabream.". Journal of Virology: JVI.01369–16. doi:10.1128/JVI.01369-16. PMID 27440877.