Extrachromosomal DNA
Extrachromosomal DNA is any DNA that is found outside of the nucleus of a cell. It is also referred to as extranuclear DNA or cytoplasmic DNA. Most DNA in an individual genome is found in chromosomes but DNA found outside of the nucleus also serves important biological functions.[1]
In prokaryotes, nonviral extrachromosomal DNA is primarily found in plasmids whereas in eukaryotes extrachromosomal DNA is primarily found in organelles.[2] Mitochondrial DNA is a main source of this extrachromosomal DNA in eukaryotes.[1] Extrachromosomal DNA is often used in research of replication because it is easy to identify and isolate.[2]
Extrachromosomal DNA was found to be structurally different from nuclear DNA. Cytoplasmic DNA is less methylated than DNA found within the nucleus. It was also confirmed that the sequences of cytoplasmic DNA was different from nuclear DNA in the same organism, showing that cytoplasmic DNAs are not simply fragments of nuclear DNA.[3]
In addition to DNA found outside of the nucleus in cells, infection of viral genomes also provides an example of extrachromosomal DNA.
Prokaryotes

Although prokaryotic organisms do not possess a membrane bound nucleus like the eukaryotes, they do contain a nucleoid region in which the main chromosome is found. Extrachromosomal DNA exists in prokaryotes outside of the nucleoid region as circular or linear plasmids. Bacterial plasmids are typically short sequences, consisting of 1 kilobase (kb) to a few hundred kb segments, and contain an origin of replication which allows the plasmid to replicate independently of the bacterial chromosome.[4] The total number of a particular plasmid within a cell is referred to as the copy number and can range from as few as two copies per cell to as many as several hundred copies per cell.[5] Circular bacterial plasmids are classified according to the special functions that the genes encoded on the plasmid provide. Fertility plasmids, or f plasmids,allow for conjugation to occur whereas resistance plasmids, or r plasmids, contain genes that convey resistance to a variety of different antibiotics such as ampicillin and tetracycline. There also exists virulence plasmids that contain the genetic elements necessary for bacteria to become pathogenic as well as degradative plasmids that harbor the genes that allow bacteria to degrade a variety of substances such as aromatic compounds and xenobiotics.[6] Bacterial plasmids can also function in pigment production, nitrogen fixation and the resistance to heavy metals in those bacteria that possess them.[7]
Naturally occurring circular plasmids can be modified to contain multiple resistance genes and several unique restriction sites, making them valuable tools as cloning vectors in biotechnology applications.[4] Circular bacterial plasmids are also the basis for the production of DNA vaccines. Plasmid DNA vaccines are genetically engineered to contain a gene which encodes for an antigen or a protein produced by a pathogenic virus, bacterium or other parasite.[8] Once delivered into the host, the products of the plasmid genes will then stimulate both the innate immune response and the adaptive immune response of the host. The plasmids are often coated with some type of adjuvant prior to delivery to enhance the immune response from the host.[9]
Linear bacterial plasmids have been identified in several species of spirochete bacteria, including members of the genus Borrelia (to which the pathogen responsible for Lyme disease belongs), several species of the gram positive soil bacteria of the genus Streptomyces, and in the gram negative species Thiobacillus versutus, a bacterium that oxidizes sulfur. The linear plasmids of prokarykotes are found either containing a hairpin loop or a covalently bonded protein attached to the telomeric ends of the DNA molecule. The adenine-thymine rich hairpin loops of the Borrelia bacteria range in size from 5 kilobase pairs (kb) to over 200 kb[10] and contain the genes responsible for producing a group of major surface proteins, or antigens, on the bacteria that allow it to evade the immune response of its infected host.[11] The linear plasmids which contain a protein that has been covalently attached to the 5’ end of the DNA strands are known as invertrons and can range in size from 9 kb to over 600 kb consisting of inverted terminal repeats.[10] The linear plasmids with a covalently attached protein may assist with bacterial conjugation and integration of the plasmids into the genome. These types of linear plasmids represent the largest class of extrachromosomal DNA as they are not only present in certain bacterial cells, but all linear extrachromosomal DNA molecules found in eukaryotic cells also take on this invertron structure with a protein attached to the 5’ end.[10][11]
Eukaryotes
Mitochondrial DNA

The mitochondria present in eukaryotic cells contain multiple copies of mitochondrial DNA referred to as mtDNA which is housed within the mitochondrial matrix.[12] In multicellular animals, including humans, the circular mtDNA chromosome contains 13 genes that encode proteins that are part of the electron transport chain and 24 genes that produce RNA necessary for the production of mitochondrial proteins; these genes are broken down into 2 rRNA genes and 22 tRNA genes.[13] The size of an animal mtDNA plasmid is roughly 16.6 kb and although it contains genes for tRNA and mRNA synthesis, proteins produced as a result of nuclear genes are still required in order for the mtDNA to replicate or for mitochondrial proteins to be translated.[14] There is only one region of the mitochondrial chromosome that does not contain a coding sequence and that is the 1 kb region known as the D-loop to which nuclear regulatory proteins bind.[13] The number of mtDNA molecules per mitochondria varies from species to species as well as between cells with different energy demands. For example, muscle and liver cells contain more copies of mtDNA per mitochondrion than blood and skin cells do.[14] Due to the proximity of the electron transport chain within the mitochondrial inner membrane and the production of reactive oxygen species (ROS), and due to the fact that the mtDNA molecule is not bound by or protected by histones, the mtDNA is more susceptible to DNA damage than nuclear DNA.[15] In cases where mtDNA damage does occur, the DNA can either be repaired via base excision repair pathways, or the damaged mtDNA molecule is destroyed (without causing damage to the mitochondrion since there are multiple copies of mtDNA per mitochondrion).[16]
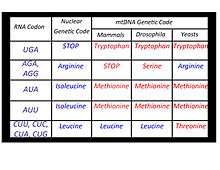
The standard genetic code by which nuclear genes are translated is universal, meaning that each 3-base sequence of DNA codes for the same amino acid regardless of what species from which the DNA comes. However, this universal nature of the code is not the case with mitochondrial DNA found in fungi and animals. While most of the 3-base sequences in the mtDNA of these organisms do code for the same amino acids as those of the nuclear genetic code, there are some mtDNA sequences that code for amino acids different from those of their nuclear DNA counterparts. Some of the coding differences found in the mtDNA sequences of mammals, fruit flies and yeasts are outlined in the adjacent table.[12] The coding differences are thought to be a result of chemical modifications in the transfer RNAs that interact with the messenger RNAs produced as a result of transcribing the mtDNA sequences.[17]
Chloroplast DNA
Eukaryotic chloroplasts, as well as the other plant plastids, also contain extrachromosomal DNA molecules. Most chloroplasts house all of their genetic material in a single ringed chromosome, however in some species there is evidence of multiple smaller ringed plasmids.[18][19][20] A recent theory that questions the current standard model of ring shaped chloroplast DNA (cpDNA), suggests that cpDNA may more commonly take a linear shape.[21] A single molecule of cpDNA can contain anywhere from 100-200 genes[22] and varies in size from species to species. The size of cpDNA in higher plants is around 120–160 kb.[12] The genes found on the cpDNA code for mRNAs that are responsible for producing necessary components of the photosynthetic pathway as well as coding for tRNAs, rRNAs, RNA polymerase subunits, and ribosomal protein subunits.[23] Like mtDNA, cpDNA is not fully autonomous and relies upon nuclear gene products for replication and production of chloroplast proteins. Chloroplasts contain multiple copies of cpDNA and the number can vary not only from species to species or cell type to cell type, but also within a single cell depending upon the age and stage of development of the cell. For example, cpDNA content in the chloroplasts of young cells, during the early stages of development where the chloroplasts are in the form of indistinct proplastids, are much higher than those present when that cell matures and expands, containing fully mature plastids.[24]
Extrachromosomal circular DNA
Extrachromosomal circular DNA (eccDNA) is present in all eukaryotic cells, is usually derived from genomic DNA, and consists of repetitive sequences of DNA found in both coding and non-coding regions of chromosomes. EccDNA can vary in size from less than 2000 base pairs to more than 20,000 base pairs.[25] In plants, eccDNA contains repeated sequences similar to those that are found in the centromeric regions of the chromosomes and in repetitive satellite DNA.[26] In animals, eccDNA molecules have been shown to contain repetitive sequences that are seen in satellite DNA, 5S ribosomal DNA and telomere DNA.[25] Certain organisms, such as yeast, rely on chromosomal DNA replication to produce eccDNA[26] whereas eccDNA formation can occur in other organisms, such as mammals, independently of the replication process.[27] The function of eccDNA has not been widely studied, but it has been proposed that the production of eccDNA elements from genomic DNA sequences adds to the plasticity of the eukaryotic genome and can influence genome stability, cell aging and the evolution of chromosomes.[28]
Viruses
Viral DNA is an example of extrachromosomal DNA. Understanding viral genomes is very important for understanding the evolution and mutation of the virus.[29] Some viruses, such as HIV and oncogenetic viruses, incorporate their own DNA into the genome of the host cell.[30] Viral genomes can be made up of single stranded DNA (ssDNA), double stranded DNA (dsDNA) and can be found in both linear and circular form.[31]
One example of infection of a virus constituting as extrachromosomal DNA is the human papillomavirus (HPV). The HPV DNA genome undergoes three distinct stages of replication: establishment, maintenance and amplification. HPV infects epithelial cells in the anogenital tract and oral cavity. Normally, HPV is detected and cleared by the immune system. The recognition of viral DNA is an important part of immune responses. For this virus to persist, the circular genome must be replicated and inherited during cell division.[32]
Recognition of viral extrachromosomal DNA by host cell
Cells can recognize foreign cytoplasmic DNA. Understanding the recognition pathways has implications towards prevention and treatment of diseases.[33] Cells have sensors that can specifically recognize viral DNA such as the Toll-like receptor (TLR) pathway.[34]
The Toll Pathway was recognized, first in insects, as a pathway that allows certain cell types to act as sensors capable of detecting a variety of bacterial or viral genomes and PAMPS (pathogen-associated molecular patterns). PAMPs are known to be potent activators of innate immune signaling. There are approximately 10 human Toll-Like Receptors (TLRs). Different TLRs in human detect different PAMPS: lipopolysacchardies by TLR4, viral dsRNA by TLR3, viral ssRNA by TLR7/TLR8, viral or bacterial unmethylated DNA by TLR9. TLR9 has evolved to detect CpG DNA commonly found in bacteria and viruses and to initiate the production of IFN (type I interferons ) and other cytokines.[34]
Inheritance
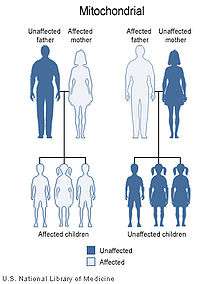
Inheritance of extrachromosomal DNA differs from the inheritance of nuclear DNA found in chromosomes. In humans, virtually all of the cytoplasm is inherited from the egg of the mother.[35] For this reason, organelle DNA, including mtDNA, is inherited from the mother. Mutations in mtDNA or other cytoplasmic DNA will also be inherited from the mother. This uniparental inheritance is an example of non-Mendelian inheritance. Plants also show uniparental mtDNA inheritance. Most plants inherit mtDNA maternally with one noted exception being the redwood Sequoia sempervirens that inherits mtDNA paternally.[36]
There are two theories why the paternal mtDNA is not transmitted to the offspring. One is simply the fact that paternal mtDNA is at such a lower concentration than the maternal mtDNA and thus it is not detectable in the offspring. A second, more complex theory, involves the digestion of the paternal mtDNA to prevent its inheritance. It is theorized that the uniparental inheritance of mtDNA, which has a high mutation rate, might be a mechanism to maintain the homoplasmy of cytoplasmic DNA.[36]
Medicine and disease
Sometimes called EEs, extrachromosomal elements, have been associated with genomic instability in eukaryotes. Small polydispersed DNAs (spcDNAs) are commonly found in conjunction with genome instability. SpcDNAs are derived from repetitive sequences such as satellite DNA, retrovirus-like DNA elements, and transposable elements in the genome. They are thought to be the products of gene rearrangements. Double minute chromosomes (DMs) are also extrachromosomal elements that are associated with genome instability.[1] DMs are commonly seen in cancer cells.[37] DMs are thought to be produced through breakages in chromosomes or overreplication of DNA in an organism. Studies show that in cases of cancer and other genomic instability, higher levels of EEs can be observed.[1]
Mitochondrial DNA can play a role in the onset of disease in a variety of ways. Point mutations in or alternative gene arrangements of mtDNA have been linked to several diseases that affect the heart, central nervous system, endocrine system, gastrointestinal tract, eye, and kidney.[13] Loss of the amount of mtDNA present in the mitochondria can lead to a whole subset of diseases known as mitochondrial depletion syndromes (MDDs) which affect the liver, central and peripheral nervous systems, smooth muscle and hearing in humans.[14] There have been mixed, and sometimes conflicting, results in studies that attempt to link mtDNA copy number to the risk of developing certain cancers. Studies have been conducted that show an association between both increased and decreased mtDNA levels and the increased risk of developing breast cancer. A positive association between increased mtDNA levels and an increased risk for developing kidney tumors has been observed but there does not appear to be a link between mtDNA levels and the development of stomach cancer.[38]
Extrachromosomal DNA is found in Apicomplexa, which is a group of protozoa. The malaria parasite (genus Plasmodium), the AIDS-related pathogen (Taxoplasma and Cryptosporidium) are both members of the Apicomplexa group. Mitochondrial DNA (mtDNA) was found in the malaria parasite.[39] There are two forms of extrachromosomal DNA found in the malaria parasites. One of these is 6-kb linear DNA and the second is 35-kb circular DNA. These DNA molecules have been researched as potential nucleotide target sites for antibiotics.[40]
Relevant research
The following research articles provide more specific details regarding extrachromosomal DNA in a variety of different organisms.
- Bogenhagen, Daniel F. (2012). "Mitochondrial DNA nucleoid structure". Biochimica et Biophysica Acta. 1819 (9–10): 914–920. doi:10.1016/j.bbagrm.2011.11.005.
- Cara, A; MS Reitz Jr (1997). "New insight on the role of extrachromosomal retroviral DNA". Leukemia. 11 (9): 1395–1399. doi:10.1038/sj.leu.2400776. PMID 9305590.
- Cohen, Sarit; Keren Yacobi; Daniel Segal (2003). "Extrachromosomal circular DNA of tandemly repeated genomic sequences in Drosophila". Genome Research. 13 (6A): 1133–1145. doi:10.1101/gr.907603. PMC 403641
 . PMID 12799349.
. PMID 12799349. - Cohen, Sarit; Marcel Mechali (2002). "Formation of extrachromosomal circles from telomeric DNA in Xenopus laevis". EMBO Reports. 3 (12): 1168–1174. doi:10.1093/embo-reports/kvf240. PMC 1308322. PMID 12446568.
- Colosimo, A.; V. Guida; G. Palka; B. Dallapiccola (2002). "Extrachromosomal genes: a powerful tool in gene targeting approaches". Gene Therapy. 9 (11): 679–682. doi:10.1038/sj.gt.3301749. PMID 12032686.
- Cummings, Donald (1979). Extrachromosomal DNA. New York: Academic Press Inc.
- Goebel, Werner (1970). "Studies on Extrachromosomal DNA Elements". European Journal of Biochemistry. 15 (2): 311–320. doi:10.1111/j.1432-1033.1970.tb01009.x. PMID 4926129.
- Jabaji-Hare, S.H.; G. Burger; L. Forget; B.F. Lang (1994). "Extrachromosomal plasmids in the plant pathogenic fungus Rhizocontia solani". Current Genetics. 25 (5): 432–431. doi:10.1007/BF00351781.
- Preer Jr., John R. (1971). "Extrachromosomal inheritance: Hereditary symbionts, mitochondria, chloroplasts". Annual Review of Genetics: 361–406. doi:10.1146/annurev.ge.05.120171.002045.
- Shibata, Yoshiyuki; P. Kumar; R. Layer; S. Wilcox; J. Gagan; J. Griffith; A. Dutta (April 6, 2012). "Extrachromosomal microDNAs and chromosomal microdeletions in normal tissues". Science. 336 (6077): 82–86. doi:10.1126/science.1213307. PMC 3703515. PMID 22403181.
- Sloan, Daniel B. (2013). "One ring to rule them all? Genome sequencing provides new insights into the 'master circle' model of plant mitochondrial DNA structure". New Phytologist. 200 (4): 978–985. doi:10.1111/nph.12395.
- Watve, Mukata M.; Neelesh Dahanukar; Milind G. Watve (February 2010). Getz, Wayne M, ed. "Sociobiological control of plasmid copy number in bacteria". PLOS ONE. 5 (2): 1–8. doi:10.1371/journal.pone.0009328.
References
- 1 2 3 4 Kuttler, Fabien; Sabine Mai (2007). "Formation of non-random extrachromosomal elements during development, differentiation and oncogenesis". Seminars in Cancer Biology. 17 (1): 56–64. doi:10.1016/j.semcancer.2006.10.007. PMID 17116402.
- 1 2 Rush, Mark; R. Misra (November 1985). "Extrachromosomal DNA in eukaryotes". Plasmid. 14 (3): 177–191. doi:10.1016/0147-619X(85)90001-0. PMID 3912782.
- ↑ Koch, J; Vogt, G; Kissel, W (May 1983). "Cytoplasmic DNA is structurally different from nuclear DNA". Die Naturwissenschaften. 70 (5): 252–4. doi:10.1007/BF00405447. PMID 6877387.
- 1 2 Nelson, David (2008). Lehninger Principles of Biochemistry. New York: W. H. Freeman and Company. pp. 307–308. ISBN 978-0-7167-7108-1.
- ↑ Watson, James (2007). Recombinant RNA: Genes and Genomes- A Short Course. New York: W. H. Freeman and Company. p. 81. ISBN 0-7167-2866-4.
- ↑ Dib, J.R.; W. Liebel; M. Wagenknecht; M. Farias; F. Meinhardt (October 2013). "Extrachromosomal genetic elements in Micrococcus". Applied Microbiology and Biotechnology. 97 (1): 63–75. doi:10.1007/s00253-012-4539-5. PMID 23138713.
- ↑ Barnum, Susan (2005). Biotechnology- An Introduction. California: Brooks / Cole. pp. 62–63. ISBN 978-0-495-11205-1.
- ↑ Laddy, Dominick; David Weiner (2006). "From plasmids to protection: A review of DNA vaccines against infectious disease". International Reviews of Immunology. 25 (3/4): 99–123. doi:10.1080/08830180600785827. PMID 16818367.
- ↑ Ongkudon, Clarence; Jenny Ho; Michael Danquah (2011). "Mitigating the looming vaccine crisis: production and delivery of plasmid-based vaccines". Critical Reviews in Biotechnology. 31 (1): 32–52. doi:10.3109/07388551.2010.483460. PMID 20879832.
- 1 2 3 Hinnenbusch, Joe; Kit Tilly (1993). "Linear plasmids and chromosomes in bacteria". Molecular Microbiology. 10 (5): 917–922. doi:10.1111/j.1365-2958.1993.tb00963.x. PMID 7934868.
- 1 2 Meinhardt, F.; R. Schaffrath; M. Larsen (1997). "Microbial linear plasmids". Applied Microbiology and Biotechnology. 47 (4): 329–336. doi:10.1007/s002530050936. PMID 9163946.
- 1 2 3 Lodish, Harvey (2013). Molecular Cell Biology, 7th edition. New York: W.H. Freeman and Company. pp. 245–251. ISBN 978-1-4641-2398-6.
- 1 2 3 Chinnery, P.F.; D.M. Turnbull (July 1999). "Mitochondrial DNA and disease". The Lancet. 354 (9176): si17–si21. doi:10.1016/S0140-6736(99)90244-1. PMID 10437851.
- 1 2 3 Dimmock, David; Lin-Ya Tang; Eric S. Schmitt; Lee-Jun Wong (July 2010). "Quantitative Evaluation of the Mitochondrial DNA Depletion Syndrome". Clinical Chemistry. 56 (7): 1119–1127. doi:10.1373/clinchem.2009.141549. PMID 20448188.
- ↑ Bohr, Vilhelm; R. Michael Anson (1999). "Mitochondrial DNA Repair Pathways". Journal of Bioenergetics and Biomembranes. 31 (4): 391–398. doi:10.1023/A:1005484004167. PMID 10665528.
- ↑ Bendich, AJ (2010). "Mitochondrial DNA, chloroplast DNA and the origins of development in eukaryotic organisms". Biology Direct. 5 (42): 42. doi:10.1186/1745-6150-5-42.
- ↑ Bernt, Matthias; Anke Braband; Bernd Schierwater; Peter Stadler (November 2013). "Genetic aspects of mitochondrial genome evolution". Molecular Phylogenetics and Evolution. 69 (2): 328–338. doi:10.1016/j.ympev.2012.10.020. PMID 23142697.
- ↑ Clegg, MT; Gaut, BS; Learn GH, Jr; Morton, BR (Jul 19, 1994). "Rates and patterns of chloroplast DNA evolution.". Proceedings of the National Academy of Sciences of the United States of America. 91 (15): 6795–801. doi:10.1073/pnas.91.15.6795. PMC 44285. PMID 8041699.
- ↑ Lilly, Jason W; Havey, Michael J; Jackson, Scott A; Jiang, Jiming (2001). "Cytogenomic analyses reveal the structural plasticity of the chloroplast genome in higher plants". The Plant Cell Online. 13 (2): 251. doi:10.1105/tpc.13.2.245. PMC 102240. PMID 11226183.
- ↑ Aronsson, Henrik; Sandelius, Anna Stina (2009). The chloroplast interactions with the environment ([Online-Ausg.]. ed.). Berlin: Springer. p. 18. ISBN 3540686967.
- ↑ Bendich, AJ (Jul 2004). "Circular chloroplast chromosomes: the grand illusion.". The Plant cell. 16 (7): 1661–6. doi:10.1105/tpc.160771. PMC 514151. PMID 15235123.
- ↑ Wang, Ying; Jun Ding; Henry Daniell; Haiyan Hu; Xiaoman Li (2012). "Motif analysis unveils the possible co-regulation of chloroplast genes and nuclear genes encoding chloroplast proteins". Plant Molecular Biology. 80 (2): 177–187. doi:10.1007/s11103-012-9938-6. PMID 22733202.
- ↑ Pfalz, Jeaneete; Thomas Pfannschmidt (April 2013). "Essential nucleoid proteins in early chloroplast development". Trends in Plant Science. 18 (4): 186–194. doi:10.1016/j.tplants.2012.11.003. PMID 23246438.
- ↑ Rowen, Beth A.; Arnold J. Bendich (2009). "The loss of DNA from chloroplasts as leaves mature: fact or artefact?". Journal of Experimental Botany. 60 (11): 3005–3010. doi:10.1093/jxb/erp158. PMID 19454766.
- 1 2 Cohen, Sarit; Andreas Houben; Daniel Segal (2008). "Extrachromosomal circular DNA derived from tandemly repeated genomic sequences in plants". The Plant Journal. 53 (6): 1027–1034. doi:10.1111/j.1365-313X.2007.03394.x. PMID 18088310.
- ↑ Cohen, Zoya; Sara Lavi (2009). Sullivan, Beth A, ed. "Replication of Independent Formation of Extrachromosomal Circular DNA in Mammalian Cell-Free System". PLOS ONE. 4 (7): 1–8. doi:10.1371/journal.pone.0006126.
- ↑ Cohen, Sarit; Neta Agmon; Olga Sobol; Daniel Segal (2010). "Extrachromosomal circles of satellite repeats and 5S ribosomal DNA in human cells". Mobile DNA. 1 (1): 1–11. doi:10.1186/1759-8753-1-11.
- ↑ Sanjuán, R; Nebot, MR; Chirico, N; Mansky, LM; Belshaw, R (October 2010). "Viral mutation rates". Journal of Virology. 84 (19): 9733–48. doi:10.1128/JVI.00694-10. PMC 2937809. PMID 20660197.
- ↑ Silverthorn, Dee Unglaub (2007). Human Physiology. Peason/Benjamin Cummings.
- ↑ "Viral Genomes".
- ↑ Lorenz, LD; Rivera Cardona, J; Lambert, PF (October 2013). Roman, Ann, ed. "Inactivation of p53 Rescues the Maintenance of High Risk HPV DNA Genomes Deficient in Expression of E6". PLOS Pathogens. 9 (10): e1003717. doi:10.1371/journal.ppat.1003717. PMC 3812038. PMID 24204267.
- ↑ Barber, GN (September 2011). "Cytoplasmic DNA innate immune pathways". Immunological reviews. 243 (1): 99–108. doi:10.1111/j.1600-065X.2011.01051.x. PMID 21884170.
- 1 2 Barber, GN (February 2011). "Innate immune DNA sensing pathways: STING, AIMII and the regulation of interferon production and inflammatory responses". Current opinion in immunology. 23 (1): 10–20. doi:10.1016/j.coi.2010.12.015. PMID 21239155.
- ↑ Griffiths, AJF (2000). An Introduction to Genetic Analysis. New York: W.H.Freeman.
- 1 2 Sato, M; Sato, K (August 2013). "Maternal inheritance of mitochondrial DNA by diverse mechanisms to eliminate paternal mitochondrial DNA.". Biochimica et Biophysica Acta. 1833 (8): 1979–84. doi:10.1016/j.bbamcr.2013.03.010. PMID 23524114.
- ↑ Shimizu, Noriaki (2011). "Molecular mechanisms of the origin of micronuclei from extrachromosomal elements". Mutagenesis. 26 (1): 119–123. doi:10.1093/mutage/geq053. PMID 21164192.
- ↑ Thyagarajan, Bharat; Renwei Wang; Heather Nelson; Helene Barcelo; Woon-Puay Koh; Jian-Min Yuan (June 2013). Bai, Yidong, ed. "Mitochondrial DNA copy number is associated with breast cancer risk". PLOS ONE. 8 (6): 1–6. doi:10.1371/journal.pone.0065968.
- ↑ Wilson, RJ; Williamson, DH (March 1997). "Extrachromosomal DNA in the Apicomplexa". Microbiology and molecular biology reviews : MMBR. 61 (1): 1–16. PMC 232597. PMID 9106361.
- ↑ Creasey, A; Mendis, K; Carlton, J; Williamson, D; Wilson, I; Carter, R (May 1994). "Maternal inheritance of extrachromosomal DNA in malaria parasites". Molecular and biochemical parasitology. 65 (1): 95–8. doi:10.1016/0166-6851(94)90118-X. PMID 7935632.