C4orf29
| Chromosome 4 open reading frame 29 | |
|---|---|
| Identifiers | |
| Symbol | C4orf29 |
| Entrez | 80167 |
| HUGO | 26111 |
| RefSeq | NP_001034806.1 |
| UniProt | Q0P651 |
| Other data | |
| Locus | Chr. 4 q28.2 |
Chromosome 4 open reading frame 29 (C4orf29) is a protein that in Homo sapiens is encoded by the C4orf29 gene.[1]
Gene
C4orf29 is found on the positive strand of the human genome at 4q28.2. It is 74.4 kbp. The gene contains 17 exons.[2] The longest mRNA transcript is composed of 13 exons and is 2200 base pairs.[1]
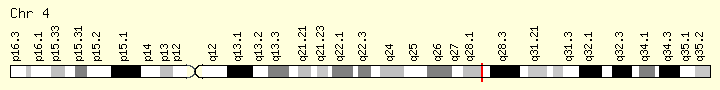
Homology
Orthologs
Many orthologs to human C4orf29 have been discovered, with the most distant ortholog with high (over 90%) coverage is found in rice Oryza sativa.[4] The protein is not found in fungi. Bacteria of the order Myxobacteria and genus Chitinimonas contain orthologous regions to the C4orf29 protein. The few bacterial homologs indicate a horizontal gene transfer event. The domain of unknown function, DUF2048, is conserved throughout orthologs.
| Organism | NCBI Accession Number | Divergence from Humans (Million Years) | Protein Length (Amino Acids) | Sequence Identity to Human | Sequence Similarity to Human |
|---|---|---|---|---|---|
| Human | NP_001034806.1 | 0 | 414 | 100% | 100% |
| Dog | XP_005631879.1 | 94.2 | 484 | 87% | 89% |
| Canary | XP_009094557.1 | 296 | 414 | 75% | 87% |
| Painted Turtle | XP_005282815.1 | 296 | 464 | 73% | 83% |
| Ant | XP_011141314.1 | 782 | 468 | 46% | 64% |
| Flat Worm | NP_492206.1 | 937 | 378 | 33% | 50% |
| Rice | NP_001046183.1 | 1369 | 366 | 31% | 47% |
| Cystobacter violaceus | WP_043404439.1 | 2535 | 321 | 31% | 52% |
Protein
C4orf29 codes a 414 amino acid sequence of 46.9 kDa in humans. The predicted isoelectric point is 9.37.[6] The domain of unknown function, DUF2048, is found from amino acid residues 25 to 414 in the precursor C4orf29 protein.[7] This domain is part of the alpha/beta hydrolase superfamily, which comprises enzymes that catalyze fat metabolism. Predicted post-translational modifications include glycosylation at residues Ser287 and Ser319 [8] and sumoylation[9] at the motifs Phe240 to Gly243, Ala377 to Asp340, and Phe408 to Gly411.
Expression
The protein product of C4orf29 in humans is predicted to be a secreted product. It is ubiquitously expressed at low to moderate levels.[10] In humans, the protein is found at high levels the digestive tract and parathyroid gland.[11] The homologous mouse protein 3110057O12Rik is expressed at high levels in the granule layer of the cerebellum.[12]
Clinical significance
C4orf29 contains highly variable numbers of Alu repeats.[13] A low number of Alu repeats in the human C4orf29 protein is associated with increase prevalence of hepatocellular carcinoma (HCC) in Asian populations. This information is used as a genetic marker to determine genetic risk of HCC.[14] Swine muscle transcriptome analysis indicates high expression of C4orf29 in swine with extreme low levels of fatty acid composition.[15]
References
- 1 2 "C4orf29". NCBI Gene. NCBI. Retrieved 5 May 2015.
- ↑ "C4orf29". NCBI AceView. Retrieved 2015-05-05.
- ↑ "C4orf29". GeneCards Human Gene Database.
- ↑ "NP_001034806". NCBI Protein BLAST.
- ↑ Kelley, LA; Sternberg, MJE. "Protein structure prediction on the web: a case study using the Phyre server". Phyre2. Retrieved 16 Apr 2015.
- ↑ "C4orf29 (human)". Phosphosite.org. Retrieved 2015-05-05.
- ↑ "uncharacterized protein C4orf29 precursor [Homo sapiens]". NCBI Protein.
- ↑ "NetOGlyc4.0 C4orf29". CBS Prediction Servers.
- ↑ "C4orf29 Sumoylation". ABGENT.
- ↑ "Large-scale analysis of the human transcriptome GDS596". NCBI GeoProfile. Retrieved 2015-05-05.
- ↑ "C4orf29". The Human Protein Atlas. Retrieved 2015-05-05.
- ↑ "3110057O12Rik". Allen Brain Atlas. Retrieved 2015-05-05.
- ↑ Osenberg, Sivan (21 Jun 2010). "Alu Sequences in Undifferentiated Human Embryonic Stem Cells Display High Levels of A-to-I RNA Editing". PLOS ONE. 5: e11173. doi:10.1371/journal.pone.0011173.
- ↑ Clifford, Robert (23 Nov 2010). "Genetic variations at loci involved in the immune response are risk factors for hepatocellular carcinoma". Hepatology. 52 (6): 2034–2043. doi:10.1002/hep.23943.
- ↑ Puig-Oliveras, A (June 2014). "Differences in muscle transcriptome among pigs phenotypically extreme for fatty acid composition". PLOS ONE. 9 (6): e99720. doi:10.1371/journal.pone.0099720. PMC 4057286
 . PMID 24926690.
. PMID 24926690.