Arcuate fasciculus
| Arcuate fasciculus | |
|---|---|
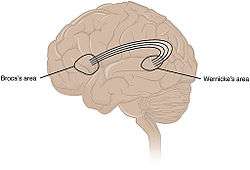 Common understanding has been that the arcuate fasciculus connects two important areas for language use, Broca's area and Wernicke's area. | |
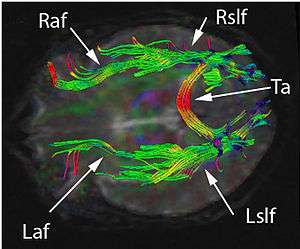 Diffusion tensor imaging image of the brain showing the right and left arcuate fasciculus (Raf & Laf). Also shown are the right and left superior longitudinal fasciculus (Rslf & Lslf), and tapetum of corpus callosum (Ta). | |
| Details | |
| Identifiers | |
| Latin | fasciculus arcuatus |
| NeuroNames | ancil-540 |
| TA | A14.1.09.557 |
| FMA | 276650 |
The arcuate fasciculus (Latin, curved bundle) is a bundle of axons that forms part of the superior longitudinal fasciculus. The arcuate bidirectionally connects caudal temporal cortex and inferior parietal cortex to locations in the frontal lobe.[1][2]
Neuroanatomy
This bundle of axons is either not present or substantially smaller in nonhuman primates.[3] Although the regions to which the arcuate fasciculus connects are still subject to debate, the connectivity of the arcuate has been shown to correspond to various functional areas within the temporal, parietal, and frontal lobes.[4] Furthermore, the topographical relationships between independent measures of white matter and gray matter integrity suggest that rich developmental or environmental interactions influence brain structure and function, and that the presence and strength of such associations may elucidate pathophysiological processes influencing systems such as language and motor planning.
Common understanding has been that the arcuate fasciculus connects two important areas for language use, Broca's area in the inferior frontal gyrus and Wernicke's area in the posterior superior temporal gyrus. As the technique of diffusion MRI has improved, this has become a testable hypothesis using brain imaging. Research indicates more diffuse termination of the fibers of the arcuate, both rostrally and caudally, than previously thought. While the main caudal source of the fiber tract appears to be posterior superior temporal cortex, the rostral terminations are mostly in premotor cortex, part of Brodmann area 6.[2][5][6]
Pathology
Evidence for the role of the arcuate fasciculus in language use is best represented by conduction aphasia, caused by damage to the inferior parietal lobule that extends into the subcortical white matter and damages the arcuate fasciculus.[7] This type of aphasia inhibits the patient from repeating unfamiliar sounds. A study by Catani, Jones, and Ffytche (2005) provided the first anatomical evidence for the presence of two pathways between Wernicke’s area and Broca’s area.[8][9] They found one deep pathway, interpreted to directly connect these two regions, and a shallower pathway that consists of two segments; the anterior segment connects frontal cortex with inferior parietal cortex, and the posterior segment connects Wernicke’s area with the inferior parietal cortex. Damage to the direct pathway may produce conduction aphasia, whereas damage to the indirect pathway spares the ability to repeat speech but impairs comprehension. The symptoms of conduction aphasia suggest that the connection between posterior temporal cortex and frontal cortex plays a vital role in short-term memory of words and speech sounds that are new or have just been heard. The arcuate fasciculus connects these two regions and circulates information back and forth, possibly contributing to short-term memory.
In nine out of ten people with tone deafness, the superior arcuate fasciculus in the right hemisphere could not be detected, suggesting a disconnection between the posterior superior temporal gyrus and the posterior inferior frontal gyrus. Researchers suggested the posterior superior temporal gyrus was the origin of the disorder.[10]
In stutterers, the arcuate fasciculus appears to have bilateral deficits that reduce it by one-third or more relative to non-stutterers.[11] However, there is ongoing debate concerning the contribution of each hemisphere and the presence of hemispheric differences, and diffusion-based evidence of differences between stutterers and controls is not isolated to the arcuate fasciculus.
See also
References
- ↑ Carlson, N. (2012). Physiology of behavior. (11th ed.). Pearson.
- 1 2 CATANI, M; THIEBAUT DE SCHOTTEN, M (1 September 2008). "A diffusion tensor imaging tractography atlas for virtual in vivo dissections". Cortex. 44 (8): 1105–1132. doi:10.1016/j.cortex.2008.05.004. Retrieved 5 October 2013.
- ↑ Rilling et. Al., 2008
- ↑ Phillips, Owen. "Topographical relationships between arcuate fasciculus connectivity and cortical thickness". Research. Human Brain Mapping. 32: 1788–1801. doi:10.1002/hbm.21147.
- ↑ Bernal, B.; Ardila, A. (18 August 2009). "The role of the arcuate fasciculus in conduction aphasia". Brain. 132 (9): 2309–2316. doi:10.1093/brain/awp206. PMID 19690094. Retrieved 5 October 2013.
- ↑ Bernal, Byron; Altman, Nolan (1 February 2010). "The connectivity of the superior longitudinal fasciculus: a tractography DTI study". Magnetic Resonance Imaging. 28 (2): 217–225. doi:10.1016/j.mri.2009.07.008. Retrieved 5 October 2013.
- ↑ Adams, R. D. The anatomy of memory mechanisms in the human brain. In "The pathology of Memory, edited by G. A. Talland and N. C. Waugh. New York: Academic Press, 1969.
- ↑ Catani, M.; Jones, D. K. (2005). "Perisylvian language networks of the human brain". Annals of Neurology. 57 (1): 8–16. doi:10.1002/ana.20319.
- ↑ Catani, M., & de Schotten, M. T. (2012). Atlas of human brain connections. Oxford University Press.
- ↑ Loui, P; Alsop, D; Schlaug, S (2009). "Tone Deafness: A New Disconnection Syndrome?". Journal of Neuroscience. 29 (33): 10215–10220. doi:10.1523/JNEUROSCI.1701-09.2009.
- ↑ Cieslak, M; Ingham, R; Ingham, J; Grafton, S (2015). "Anomalous white matter morphology in adults who stutter". Journal of Speech, Language, and Hearing Research. 58: 268. doi:10.1044/2015_JSLHR-S-14-0193.
External links
| Wikimedia Commons has media related to Arcuate fasciculus. |
- https://web.archive.org/web/20070528184714/http://www.lib.mcg.edu:80/edu/eshuphysio/program/section8/8ch15/s8c15_14.htm
- http://thebrain.mcgill.ca/flash/d/d_10/d_10_cr/d_10_cr_lan/d_10_cr_lan.html
- http://brain.oxfordjournals.org/cgi/reprint/awh622v1.pdf